
Ключевые слова: север, биота, климат, индексы тепла и сухости, таксоны, взаимосвязи
Введение. К северу Тюменской области относится территория Ямало-Ненецкого (ЯНАО) и Ханты-Мансийского (ХМАО) автономных округов общей площадью 1304 км2(это значительно больше Германии, Франции и Испаниивместе взятых) и протяженностью в меридиональном направлении более 1500 км. Это главная кладовая углеводородного сырья России, в ее недрах содержится 91% разведанных запасов газа и 46% нефти.
Общим для региона являются суровые природные условия, определяющие здесь, особенно в его арктической части, скудость биологических ресурсов и трудности хозяйственного освоения. Онвключает в себя восемь биоклиматических комплексов (БК). Их наименования и нумерация (I, II, …VIII) приведены на рис.1. Биотическому богатству и разнообразию региона, их зависимости от климата посвящена обширная литература [5, 12, 14],и др. В то же время недостаточно отражены данные о связи климата с различными показателями биоты (всевозможными формами влияния организмов друг на друга и на среду), особенности их распределения в пространстве и времени и связи с определенными температурными интервалами. Целью данной статьи является устранение этих недостатков. Материал распределен по блокам: климатическому и биотическому. В первом анализируются взаимосвязи основных элементов климата (ЭК), во втором – закономерности зависимости биоты, ее флористической и фаунистической составляющей, от ключевых климатических характеристик. Климатические показатели взяты по данным метеостанций. Аппроксимации искомых зависимостей и их достоверность (коэффициент детерминацииR2 ) определялись по программе Excel.
Основные климатические показатели и их взаимосвязи
Важнейшими комплексными показателями климата являются: индекс сухости J = В/Urг (В - годовой радиационный баланс, ккал/см2; U =0,6 ккал/см3– теплота испарения, rг – годовая сумма осадков, см), характеризующий соотношение поступления в почву тепла и влаги, и суммы положительных S>0 и отрицательных S< 0 температур воздуха, градусосутки (гс) – индексы тепла и холода, ответственные за теплообмен у поверхности Земли [2]. Входящее в J отношение В/U= rm – это метрический эквивалент радиационного баланса, характеризующий максимально возможное испарение – испаряемость, а индекс сухости J = В/Urг = rm/ rг есть количественный критерий, указывающий на избыток (или недостаток) тепла или влаги. Если J<1, то в избытке влага, если J>1, – тепло. Соответственно, в первом случае жизнь биоты и ее эволюция зависят, в первую очередь, от поступления тепла, во втором – влаги. Таким образом, изолиния J ≈1 делит биосферу на северную (холодную, влажную) и южную (теплую, сухую). Автономные округа находятся в северной фитосфере, характеризующейся возрастанием обилия и разнообразия биоты с севера на юг (в южной фитосфере – наоборот [2]). В агрономии соотношение между теплом и влагой определяется гидротермическим коэффициентом Селянинова К.о = rт / ∑>0, где rт - сумма осадков (см) за теплое время года [15].
На рис.1 – 2 приведены схематические карты [8] биоклиматического районирования севера Тюменской области и распределения определяющих ЭК.
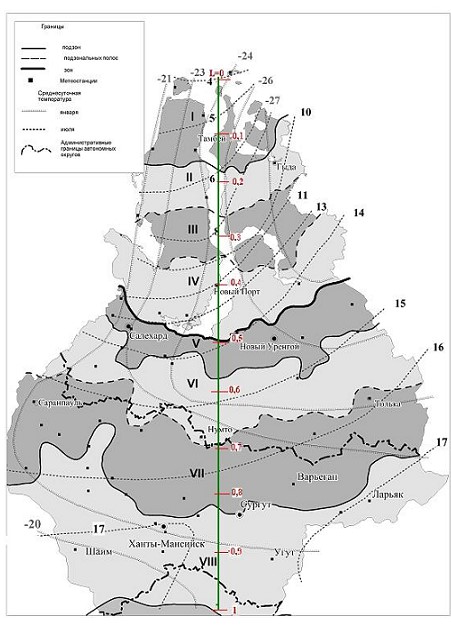
Рис. 1. Биоклиматические комплексы – БК[11]
(I– арктическая тундра, IIи III– северная и южная полоса субарктических типичных тундр, IV– субарктические кустарниковые тундры, V- лесотундра, VIи VII- северная и южная полоса северной тайги, VIII– средняя тайга; среднемесячные температуры воздуха января и июля на севере Тюменской обл. Вертикаль L=1 – условный меридиан, может служить масштабной шкалой с ценой деления ~150 км при определении расстояний между изолиниями ЭК.)
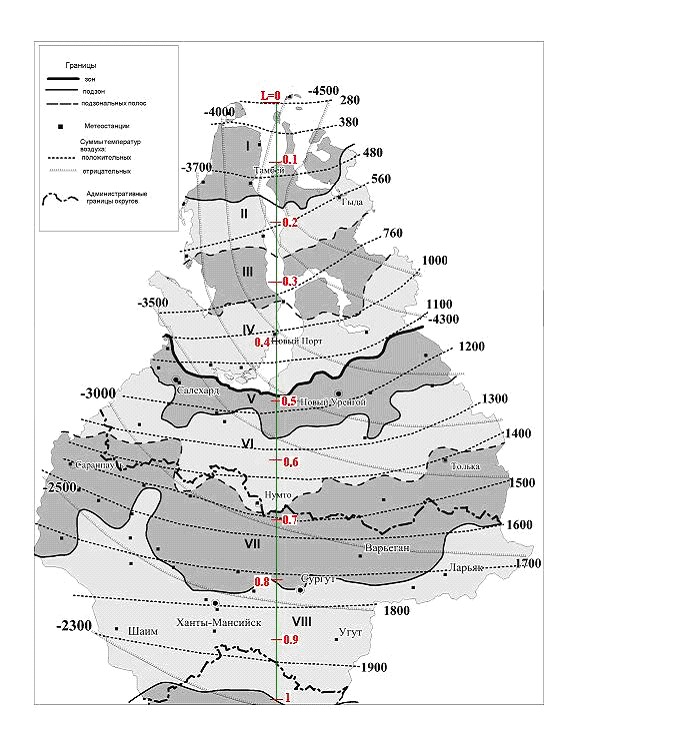
Рис. 2. Суммы положительных и отрицательных температур (градусосутки, гс) на севере Тюменской области [11]
(I– арктическая тундра, IIи III– северная и южная полоса субарктических типичных тундр, IV– субарктические кустарниковые тундры, V- лесотундра, VIи VII- северная и южная полоса северной тайги, VIII– средняя тайга; среднемесячные температуры воздуха января и июля на севере Тюменской обл. Вертикаль L=1 – условный меридиан, может служить масштабной шкалой с ценой деления ~150 км при определении расстояний между изолиниями ЭК.)
Все элементы климата (ЭК), как единой системы, взаимосвязаны. На рис. 3 приведены графики, связывающие наиболее значимые для биоты ЭК на севере Тюменской области и их аппроксимации; кроме уже известных по тексту обозначений, здесь: S>5 и S>10 – суммы температур выше 5 и 10оС; jrт = rт/ rг – относительная величина суммы осадков за теплый период; Аm= tm +(-t1) – амплитуда колебаний температуры воздуха, tm и t1 – среднемесячные максимальная (на побережье и островах – август tm= t8, южнее – июль tm= t7,) и минимальные температуры воздуха.
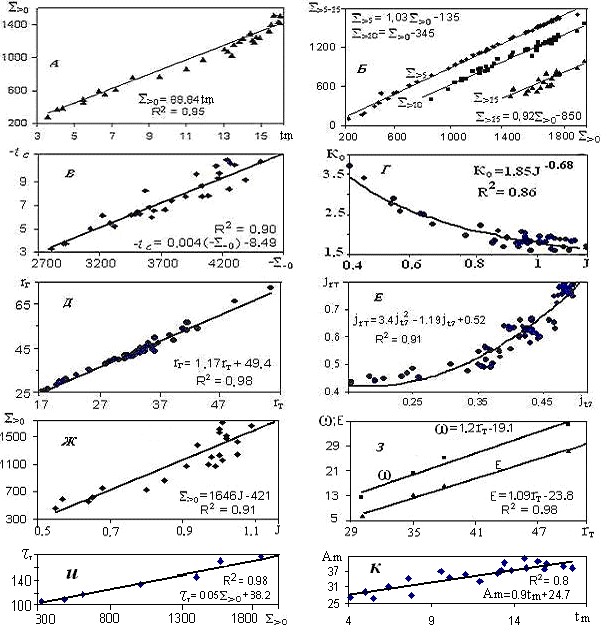
Рис. 3. Графики зависимостей:S>0 от t7– А; S> (5-15)от S> 0 – Б; tcот S- 0– В; Коот J– Г; rгот rт – Д; jrтот jt7 – Е; S>0 от J– Ж; ω и Е от rТ– З;τтот S>0 – и; Аmот tm – К(обозначения в тексте).
Территория округов – область многолетнего (ЯНАО) и сезонного (ХМАО) промерзания грунтов
В тундре и лесотундре многолетнемерзлые породы имеют сплошное распространение, их мощность увеличивается от 250-300 м на широте Полярного круга до 500-550 м на 70ос.ш. Южная граница сплошной многолетней мерзлоты примерно совпадает с изолинией S> 0 = 1200 гс. Глубина сезонного оттаивания увеличивается с севера на юг, от 1.3 до 2.7 м у песчаных грунтов, и от 0.2 до 0.5 м у торфяных. На рис. 4 показано распределение максимальной глубины сезонного оттаивания hт песчаных грунтов, рассчитанной по известной формуле Стефана [6], отображающей квадратичную зависимость глубины оттаивания от времени или индекса тепла: hт ~ τ 0.5 ~ (S>0)0.5 ..
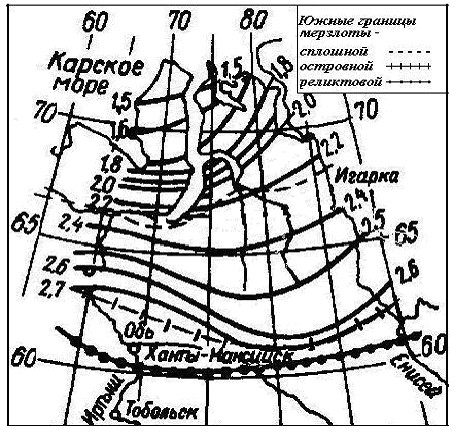
Рис. 4. Распределение глубины сезонного оттаивания hт песчаных грунтов [11]
Ход глубины сезонного оттаивания hτ в теплый период года τт в конкретном месте зависит от текущих значений времени τ или индекса тепла (S>0)τ .
hτ = hт ∙ (τ / τт)0.5 = hт ∙ [(S>0)τ /S>0] 0.5 , (1)
Для перехода к другим грунтам значение hт, найденное по рис.4, умножается на понижающий коэффициент, примерно равный: 0.8 для глинистых грунтов, 0.65 для суглинистых и 0.4 для торфяных.
В теплое время года подошва оттаивающего слоя служит водоупором, вызывающим его переувлажнение. Таяние мерзлоты и недостаточное испарение способствуют заболачиванию территории и развитию здесь специфического озерно-болотного ландшафта с преобладанием травянистой растительности. В период таяния мерзлого слоя из него идет интенсивное выделение защемленных газов, преимущественно метана и диоксида углерода – главных виновников парникового эффекта и повышенной пожарной опасности.
О подобии климатических и пространственно-временных показателей
В пределах холодной фитосферы изменение климатических показателей в течение теплого периода года, в частности среднемесячных температур и их сумм, подобно их пространственному изменению в направлении с севера на юг. Влажность почвы здесь избыточна, поэтому биота зависит, в основном, от атмосферного тепла, опосредованного величиной S>0. Чем выше S>0 (и tм), тем через большее количество природных зон, находящихся севернее арктической пустыни, где S>0 близка к 0, дважды (туда к середине лета и обратно к началу зимы) «пробегает» данное географическое место в течение теплого периода года, и тем обильнее и разнообразней его биота. Изменение индекса тепла в теплое время года на равнинной местности подобно изменению этого параметра в меридиональном направлении (с севера на юг). В горах аналогичным пространственным фактором является высотная поясность, когда индекс тепла убывает с повышением высоты. Индекс тепла как время или пройденный путь – кумулятивные величины, изменяющиеся только в сторону увеличения.
На рис. 5 показан многолетний ход годового индекса тепла в Сургуте (а), Салехарде (б) и Березове (в), а на рис. 5 г – ход его суммарного роста в тех же пунктах, но в относительных (нормализованных) величинах j. Все три кривые на рис. 5 г практически сливаются. Графики на рис. 5 д отражают зависимость годового индекса тепла в размерной и безразмерной форме от jL= ΔL/L – относительного расстояния по линии L на рис.1. Из сравнения графиков на рис. 5 г и д следует примерная идентичность (эквивалентность, взаимозаменяемость) всех трех нормализованных величин – индекса тепла, времени и пространства. Т.е. в первом приближении, для всего региона справедливо равенство jS>0 = jτ = jL .
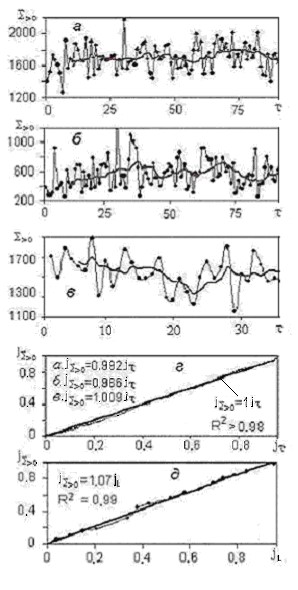
Рис. 5. Ход S>0 (гс) во времени τ (годы):
а – Сургут, б – Салехард, в -Березово; зависимость jS>0 от jτ – г и от jL – д
Климатическая зависимость биотического богатства и разнообразия
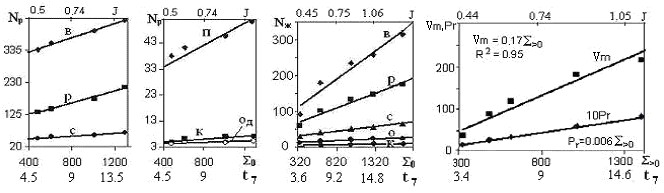
Рис. 6. Графики зависимости Nж,Nр, Vmи Prот S>0, а также от t7 (вторая нижняя горизонтальные ось) иJ(верхняя горизонтальная ось) на севере Тюменской области
На графиках рис. 6 показана зависимость количества таксонов разного уровня: видов (В), родов (Р), семейств (С), порядков (П) или отрядов (О), классов (К) и отделов (Од) сосудистых растений (Nр) [8] и животных (Nж ) [7], а также продуктивности (годичной первичной продукции) Рr, т/(га ∙ год) и фитомассы Vm, т/га [1]. от климатических показателей: S>0,t7 и J.
Анализ таблиц и графиков зависимости биотических показателей от индекса тепла показал, что:
а) начиная с уровня классов количество таксонов перестает зависеть от климата, становится примерно постоянным, одинаковым для всех БК;
б) общая формула зависимости параметров биоты от S>0 практически линейна:
У=АS>0+В (2)
где У – общее обозначение биотических показателей, А и В – численные коэффициенты, определяемые по табл.1.
Формула (2) вкупе с табл. 1 позволяют определить количество биотических таксонов непосредственно по климатическим показателям – S>0, t7 или J.
Таким образом, количество таксонов любого уровня, примерно до класса, зависит от климата, увеличиваясь с севера на юг. В то же время, как следует из рис.7 (а и б), отношение количества таксонов любого уровня к количеству видов – постоянная величина, не зависящая от климата (т.е. во всех природных комплексах Nр2/ Nр1=0,44; Nж2/ Nж1=0,58 и т.п.). Поэтому, зная количество видов, по формулам на рис. 7 можно рассчитать и количество таксонов более высокого ранга (родов, семейств). Рис.7в отражает устойчивую линейную связь таксонов флоры и фауны, также инвариантную климату.
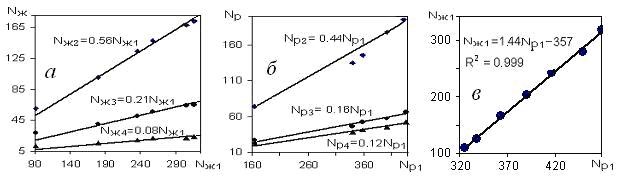
Рис. 7. Зависимость численности таксонов животных Nж2 - Nж4 от Nж1 и Nр2 - Nр4 от Nр1 (а, б), а такжеNж1 от Nр1(в); (1 – вид, 2 – род, 3 – семейство, 4 – отряд)
Таблица 1 - Постоянные в формуле (1) для основных параметров групп биоты (ГБ): количества таксонов птиц (Пт), млекопитающих (М), всех животных Nж=М + Пт, древесных (Д) и травянистых (Тр) растений, всей растительности (Nр), продуктивности Pr и биомассы Bm растений, а также значения R2
ГБ | Таксоны | А | В | R2 | ГБ | Таксоны | А | В | R2 |
Пт | Виды | 0.125 | 53.1 | 0.92 | Д | виды | 0.0024 | 55 | 0.88 |
Роды | 0.063 | 34.9 | 0.93 | роды | 0.001 | 26 | 0.87 | ||
cем-cтва | 0.02 | 16.1 | 0.94 | cем-cтва | 0.0003 | 14.3 | 0.73 | ||
отряды | 0.008 | 5.8 | 0.9 | Тр | виды | 0.09 | 255 | 0.99 | |
М | Виды | 0.031 | 10.3 | 0.98 | роды | 0.09 | 60 | 0.97 | |
Роды | 0.017 | 10.1 | 0.95 | cем-cтва | 0.02 | 25 | 0.94 | ||
cем-cтва | 0.006 | 6.8 | 0.96 | Nр | виды | 0.115 | 285 | 0.98 | |
отряды | 0.001 | 4.5 | 0.80 | роды | 0.096 | 86.3 | 0.94 | ||
Nж | Виды | 0.176 | 49.8 | 0.98 | cем-cтва | 0.022 | 36.4 | 0.91 | |
Роды | 0.091 | 37.2 | 0.98 | порядки | 0.021 | 25.4 | 0.9 | ||
cем-cтва | 0.029 | 21 | 0.98 | классы | 0.003 | 3.6 | 0.86 | ||
Отряды | 0.01 | 9.2 | 0.99 | отделы | 0.0026 | 2.7 | 0.84 | ||
Pr | – | 0.006 | 0 | 0.98 | Vm | – | 0.17 | 0 | 0.96 |
Температурные интервалы и аффилированные с ними биотические группы
Биотическое разнообразие выражают через различные соотношения между массой или численностью разных групп биоты (совокупность популяций, населяющих определенную территорию, которая функционирует как единое целое благодаря взаимосвязанным метаболическим превращениям) – индексы Шеннона (мера энтропии), Симпсона (мера дисперсии) и др. [4]. Чем больше индекс Шеннона, тем разнообразней биота. Увеличение индекса Симпсона соответствует росту доминирования. Следует отметить, что состав биоты, количество ее систематических групп (таксонов) и соотношения между ними реально определяются только непосредственным подсчетом (переписью) в полевых условиях, т.е. все известные индексы разнообразия биоты рассчитываются по уже установленным ее показателям. Причем ни один из известных индексов не отражает влияние климатических факторов. Между тем именно климат, определяющий тепло- и влагообеспеченность, является главным (первичным) фактором членения биоты, ее многообразия. Он позволяет оценивать, хотя бы приближенно, структуру биоты только по климатическим данным, в частности, по индексам тепла S>0 и сухости или по максимальным среднемесячным температурам tм.
В климатических справочниках, помимо ∑>0, даются суммы температур выше 5, 10 и 15оС, которые хорошо коррелируют с ∑>0 – рис.3. Выделим четыре температурных интервала с разными тепловыми условиями вегетации и соответственно структурой растительности: 1) Δt = 0-5; 2) Δt =5-10; 3) Δt =10-15 и 4) Δt =15-tmоС и распределим между ними сумму положительных температур ∑>0 – условный показатель тепла. Каждому интервалу, а фактически определенному климатическому поясу – 1) арктическому, 2) субарктическому, 3) умеренному и 4) засушливому отвечает определенная группа биоты, существующая при этих температурах: 1) арктическая (Ар), 2) субарктическая или морозостойкая (Мс), 3) теплолюбивая (Тл), 4) сухоустойчивая (Су). В средней тайге и южнее, где t7 > 15оС, распределенное условное тепло вычисляется по формулам: 1) ∑1 = ∑>0 -∑>5 ; 2) ∑2= ∑>5 -∑>10 ; 3) ∑3= ∑>10 -∑>15 . Последний (в данном случае четвертый) интервал определяется по остаточному принципу: 4) ∑4 = ∑>0 – (∑1 +∑2 + ∑3) = ∑>15. Такое групповое структурирование условного тепла и обусловленных им биотических показателей объясняет (наряду с историей климата) часто наблюдаемое распространение определенных видов биоты далеко за пределами своего ареала, наличие категории редких видов и ряд других особенностей распределения биоты.
Выделение тепловых интервалов продемонстрируем на примере Сургута, где ∑>0 =1734, ∑>5 =1644, ∑>10 = 1361, ∑>15 =791, t7= 16.9оС, τт = 180 сут, hт = 2.7м. Тогда: ∑1 = 90 гс; ∑ 2 = 283 гс; ∑ 3 = 570 гс; ∑4 =1734 – (90 + 283 +570) = ∑>15 =791 гс. Доли (η1-4 = ∑1-4 /∑>0) от всей суммы положительных температур в каждом интервале равны: η1=90/1734=0,05; η2=283/1734=0,16; η3=570/1734=0,33 и η4=791/1734=0,46.
При анализе нужно различать полную (максимальную) сумму положительных температур ∑>0 и изменяющуюся (растущую) в течение теплого периода (∑>0)τ от 0 до ∑>0 (в Сургуте от 0 до 1734 гс). Ход (τ, час) температур воздуха t (оС) и их сумм (∑>0) τ в теплое время года в Сургуте показаны на рис. 8. Оба графика с высокой достоверностью (R2 ≈ 0,99) описываются полиномом:
(∑>0)τ; t= aτ2+bτ (3)
При расчете (∑>0)τ a=0. 01, b=9; при расчете t b= -0.002 , b=0.37
Ход температур воздуха tв теплое время хорошо описывается также синусоидой, а их сумм (∑>0)τ– линейной функцией:
t≈ t0+ tmsin(πτ/τт), (∑>0)τ ≈ ∑>0 ∙ τ/τт (4)
где t0– температура начала (и конца) теплого времени года, в умеренных и северных широтах t0 ≈ 0 оС; τ - текущее время теплого периода; τт – длительность теплого периода.
Величину (∑>0)τ можно определить и как интеграл синусоиды (5):
(∑>0)τ = tm τт/3.14 [1-cos(3.14 τ/τт)] (5)
На рис. 8 выделены все четыре интервала температур, соответствующие им индексы тепла и группы биоты снизу вверх: арктическая (Ар), морозоустойчивая (Му), теплолюбивая (Тл), сухоустойчивая (Су).
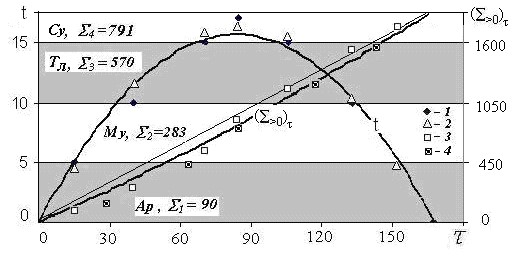
Рис. 8. Ход температур воздуха t(оС), их сумм (∑>0)τ(гс) в теплое время года (τ, сут), в Сургуте и их аппроксимации : 1-синусоидальняя, 2- полиномиальная, 3 – линейная, 4 - косинусоидальняя.
Севернее, где среднемесячные температуры воздуха ниже 15оС, 4-й участок отсутствует; величины распределенного тепла на остальных участках определяются аналогично, в том числе на 3-м (последнем): ∑3=∑10-t7 = ∑>0 – (∑0-5 +∑5-10)= ∑>10. По такой же схеме составляются формулы для расчета распределенного тепла на Крайнем Севере, где отсутствует третий, а при tm≤ 5oCи второй интервалы. Например, в Тамбее, где ∑>0 =493, ∑>5 =344 гс, а самая высокая среднемесячная температура летом tм= t8= 6.4>5оС, третий и четвертый участки отсутствуют.Отнимая ∑>5 =344 от ∑>0 =493, получаем количество условного тепла на первом температурном участке ∑1= ∑0-5 = 149 гс; остальное относится ко второму участку ∑2=∑>5=344 гс. Групповые ∑1-nтепловые индексы – константы для каждого географического места (n– число интервалов: от 1 в арктической пустыне до 4 в тайге) можно использовать для оценки богатства и разнообразия биоты.
На уровне групповых индексов тепла также наблюдается подобие(эквивалентность) временных и пространственных (в данном случае вертикальных) страт. Под последними понимаются значения hτ, фиксирующие нижние границы отдельных слоев сезонного оттаивания, отвечающих за вегетацию выделенных выше 4-х групп биоты. Они рассчитываются по формуле (1), при этом входящие в неё (∑>0)τили τ определяются с помощью формул (3) – (5) либо снимаются непосредственно с графиков, типа показанного на рис. 8. Для примера в табл. 2 приведены выделенные группы биоты Ар, Му, Тл и Су и определяющих ихзначения τ, (∑>0)τи hτдля Сургута согласно рис. 8. Во втором столбце таблицы также показаны их величины (нули), отвечающие состоянию дневной поверхности на начало весны.
Таблица 2 - Выделенные группы биоты (ГБ) и определяющие их параметры: τ, сутки; (∑>0)τ, гс; hτ, м.
ГБ | - | Ар | Му | Тл | Су |
τ | 0 | 55 | 95 | 150 | 180 |
(∑>0)τ | 0 | 450 | 1050 | 1600 | 1734 |
hτ | 0 | 1.4 | 2.1 | 2.6 | 2.7 |
Анализ показал, что зависимость биотических таксонов от индекса тепла с несколько большей, но также допустимой погрешностью (0.8-0.9) можно аппроксимировать упрощеннойформулой (2), при В=0, когда все параметры биоты (не только Prи Vm, как в табл. 1) и суммы температур прямо пропорциональны. При этом коэффициент пропорциональности равенη1- n, а величина А определяется по табл. 3. Это позволяет вычислять все биотические показатели, в том числе и распределенные по температурным интервалам (У1- n) по общей формуле:
У1- n= У · η1- n (6)
Таблица 3 - Коэффициенты Ав упрощенной формуле (1) для основных параметров групп биоты (ГБ): количества таксонов птиц (Пт), млекопитающих (М), всех животных Nж=М + Пт, древесных (Д) и травянистых (Тр) растений, всех сосудистых растений (Nр = Д+Тр).
ГБ | Пт | М | Nж | Д | Тр | Nр |
А | 0.56 | 0.04 | 0.21 | 0.19 | 1.3 | 0.2 |
Итоговым результатом работы является табл. 4, в которую сведены основные ЭК, осредненные по выделенным биоклиматическим комплексам (БК) и зависящее от них видовое богатство и разнообразие биоты, общее и распределенное по температурным интервалам.
Из табл. 4 следует, что арктическая биота (Nр1,. Nж1) встречается по всему региону, убывая с севера на юг. Это соответствует фактическим данным. Например, пыльца карликовой березки – эндемика тундры – повсеместно присутствует и в таежных поверхностных палиноспектрах [9]. Сухоустойчивая биота (Nр4,. Nж4) встречается только в таежной зоне и южнее. Распределение численности субарктических (N2) и теплолюбивых (N3) видов имеет более сложный волнообразную форму. Причем максимальные значения долевых тепловых индексов и соответствующие им численности видов биоты (и растений и животных) приурочены к биокомплексам III(юг типичной тундры) и VI(север северной тайги).
В табл. 4 приведены только количества видов распределенных по температурам растений и животных. При необходимости по формулам на рис.6 нетрудно определить эти показатели и для последующих иерархических уровней (родов, семейств и т.д.).
Таблица 4 - Максимальные среднемесячные температуры (tm,оС); индексы сухости (J) и тепла (∑>0), гс; число видов растений (Nр) и животных (Nж) в разных БК (по рис.1). А также долевые тепловые индексы η1-4 и распределенные по четырем температурным интервалам числа видов растений (Nр1-4) и животных (Nж.1-4), синим выделены их максимумы
БК | I | II | III | IV | V | VI | VII | VIII |
tm | 5,3 | 6 | 9 | 12,5 | 14 | 15 | 16 | 18 |
J | 0,45 | 0,5 | 0,6 | 0,7 | 0,75 | 0,81 | 0,88 | 0,96 |
∑>0 | 460 | 610 | 760 | 1050 | 1230 | 1320 | 1600 | 1850 |
η1 | 0,27 | 0,19 | 0,15 | 0,1 | 0,08 | 0,07 | 0,05 | 0,04 |
η2 | 0,72 | 0,81 | 0,85 | 0,23 | 0,21 | 0,19 | 0,16 | 0,14 |
η3 | – | – | – | 0,67 | 0,72 | 0,73 | 0,31 | 0,27 |
η4 | – | – | – | – | – | – | 0,47 | 0,55 |
Nр | 326 | 338 | 364 | 390 | 410 | 457 | 470 | 480 |
Nр1 | 90 | 64 | 55 | 39 | 36 | 31 | 24 | 19 |
Nр2 | 236 | 274 | 309 | 90 | 95 | 98 | 84 | 67 |
Nр3 | – | – | – | 261 | 280 | 328 | 145 | 130 |
Nр4 | – | – | – | – | – | – | 220 | 264 |
Nж | 115 | 127 | 166 | 204 | 233 | 280 | 316 | 323 |
Nж1 | 31 | 24 | 25 | 20 | 20 | 20 | 16 | 13 |
Nж2 | 84 | 103 | 141 | 47 | 51 | 53 | 51 | 45 |
Nж3 | – | – | – | 137 | 161 | 207 | 98 | 87 |
Nж4 | – | – | – | – | – | – | 151 | 178 |
Потепление климата и его влияние на биоту
В последние 40-50 лет отмечается повсеместное потепление климата, основным признаком которого является повышение температуры воздуха, как в теплое время года, так и в среднегодовом выражении. Потепление вызывает много негативных последствий, часто катастрофического характера – пожары, наводнения, просадки вечномерзлых грунтов, разрушительные деформации инженерных сооружений и т.п. Особенно оно опасно для Севера, где грозит оттаиванием приполярных и подземных льдов и высвобождением огромных масс воды и защемленных в толще мерзлоты газов. В то же время, очевидно, что повышение температур воздуха играет и положительную роль, увеличивая длительность вегетационного периода и продуктивность биоты. Оценим эту роль для севера Тюменской области, используя климатические справочники 1965 и 2011 годов.
В табл. 5 приведены среднемноголетние значения среднегодовых (tc) и максимальных среднемесячных (tm) температур воздуха в ряде пунктов на севере Тюменской обл. за периоды до 2011 и до 1965 гг.
Таблица 5 - Средние значения tcиtmза периоды времени до 2011 (а) и 1965г.(b) и их изменения за 50 лет на севере Тюменской области.
№ | Пункт | tc..а , 2011 | tс.b , 1965 | tm.а, 2011 | tm.b, 1965 | tm.а/tm.b/ |
1. |
о.Белый |
-11.7 | -10.4 |
4.9 |
4.1 |
1.19 |
2. | Харасавэй | -10.5 | -9.8 | 6.6 | 5.5 | 1.2 |
3. | Тазовский | -8.6 | -9.3 | 14.5 | 13.4 | 1.08 |
4. | Сидоровск | -8 | -8.5 | 15.6 | 14.6 | 1.07 |
5. | Н.Порт | -7.8 | -9.4 | 12.2 | 11 | 1.11 |
6. | Ямбург | -6.3 | -6.9 | 14.3 | 13 | 1.1 |
7. | Салехард | -6.3 | -6.4 | 14.7 | 13.8 | 1.06 |
8. | Халесавэй | -5.3 | -5.8 | 17.2 | 15.9 | 1.08 |
9. | Тарко-Сале | -6 | -6.7 | 16.4 | 15.4 | 1.06 |
10. | Яр-Сале | -7.3 | -7.5 | 14.4 | 13.2 | 1.09 |
11. | Надым | -5.9 | -6.6 | 15.9 | 14.7 | 1.08 |
12. | Березово | -3.1 | -3.8 | 16.4 | 15.8 | 1.04 |
13. | Сургут | -2.9 | -3.1 | 17.5 | 16.9 | 1.04 |
14. | Няксимволь | -2.2 | -1.2 | 17.3 | 15.8 | 1.09 |
15. | Х.-Мансийск | -0.8 | -1.4 | 18.3 | 17.5 | 1.05 |
Табл. 5 отражает общую тенденцию к повышению максимальных среднемесячных температур воздуха в последние примерно 50 лет: на 0,6 – 1,5оС (в среднем примерно на 1оС) или на 5-19% со средней скоростью: vtm≈ 1/50 ≈ 0,02 град/год. Причем наибольшее повышение в процентном отношении наблюдается на севере региона, к югу оно убывает. Что касается среднегодовых температур, то на большей части территории они также повышаются, за исключением островов и побережья Карского моря (о. Белый и м. Харасавэй), где наблюдается некоторое их понижение. Это говорит о том, что потепление происходит, в основном за счет повышения летних температур.
Увеличение индекса тепла вызывает перемещение биоклиматических комплексов (БК) с юга на север и увеличение (в целом) видового богатства биоты. В то же время увеличение максимальной среднемесячной температуры воздуха на один градус на побережье вызовет потерю части ареала арктической наземнойбиоты, соответственно ее уменьшение и переход в категорию редких видов - объектов Красной книги.
В табл. 6 приведены значения индекса тепла за периоды наблюдений до 1965 г. (числители) и до 2011 г. (знаменатели), взятые из справочников и соответствующие им величины видового богатства и продуктивности, рассчитанные по формуле (2). Аналогично можно оценить влияние потепления на групповые индексы тепла, биомассу и биотические таксоны высших рангов.
Таблица 6 - Индексы тепла (∑>0), гс; продукция (Pr, т/га·год); число видов растений (Nр) и животных (Nж) в разных БК (по рис.1), соответствующие климатическим показателям наблюдаемым до 1965 г (числитель) и до 2011 г. (знаменатель).
БК | I | II | III | IV | V | VI | VII | VIII |
∑>0 | 340/432 | 439/531 | 658/750 | 877/969 | 1097/1189 | 1316/1408 | 1536/1628 | 1700/1819 |
Pr | 2/2,2 | 2,6/2,8 | 3,9/4,2 | 5,3/5,6 | 6,6/7 | 7,9/8,4 | 9,2/9,9 | 10,2/10,9 |
Nр | 327/335 | 338/346 | 364/371 | 390/396 | 417/421 | 449/457 | 469/472 | 480/491 |
Nж | 110/125 | 127/143 | 166/181 | 204/220 | 243/259 | 281/298 | 316/336 | 324/365 |
Повышение максимальных среднемесячных температур воздуха за последние примерно 50 лет на 1оС, согласно табл. 6, должно вызвать увеличение видов: сосудистых растений от 8 арктической тундре до 11 в средней тайге; животных от 15 в арктической тундре до 41 в средней тайге.
Заключение
1. Основными климатическими показателями, определяющими богатство и разнообразие биоты, являются индексы сухости и тепла. Количество биотических таксонов в пределах севера Тюменской области увеличивается с севера на юг вслед за увеличением этих показателей. В то же время, отношение количества родов, видов, семейств, отрядов (порядков), к количеству видов остается постоянным. Количество видов флоры и фауны устойчиво увязано друг с другом, а их отношение также инвариантно климату.
2. За последние 50 лет июльские температуры воздуха на севере Тюменской области повысились примерно на 1оС, а суммы положительных температур – на 93 гс. Такое потепление соответствует увеличению количества видов: растений на 1-3%, животных в – среднем на 8-9%. Т.е., позитивное влияние потепления климата на биоту севера имеет место, но в общем, оно невелико и вряд ли компенсирует связанные с потеплением негативные факторы – эмиссию газов из оттаивающих льдов, пожары, наводнения и т.п.
Список литературы:
2. Будыко М.И. Климат и жизнь. Л., Гидрометеоиздат. 1981. 400 с
3. Гашев С.Н. Млекопитающие Тюменской области: справочник-определитель. Тюмень: Изд-во ТюмГУ, 2008. 333 с.
4. Гелашвили Д.Б., Иудин Д.И., Розенберг Г.С. и др. Основы мультифрактального анализа видовой структуры сообщества // Успехи современной биологии. 2008, т. 128. С.. 21-34
5. Ильина И.С., Лапшина Е.И., Лавренко Н.Н. и др. Растительный покров Западно- Сибирской равнины. Новосибирск: Наука, 1985. 250 с.
6. Коновалов А.А., Роман Л.Т. Особенности проектирования фундаментов в нефтепромысловых районах Западной Сибири. Л.: Стройиздат. 1981. 168 с.
7. Коновалов А.А., Гашев С.Н., Казанцева М.Н. Распределение и иерархия биотических таксонов на территории Тюменской области // Аграрная Россия. 2013, 4. С. 48-57.
8. Коновалов А.А., Глазунов В.А. Московченко Д.В. Тигеев А.А. Гашев С.Н. О климатической зависимости биоты на севере Тюменской области. Материалы IIIВсероссийского совещания «Человек и Север». Тюмень: Изд-во ИПОС СО РАН, 2015. С. 333-338.
9. Коновалов А.А., Иванов С.Н. Климат, фитопродуктивность и палиноспектры: связи, распределение и методика палеореконструкции (на примере Западной Сибири. Новосибирск: ГЕО, 2007. 130 с.
10. Коновалов А.А. Климатическая зависимость биоты (на примере Западной Сибири). ЖОБ, т.79, №1, 2018, с. 55-63.
11. Научно-прикладной справочник «Климат России, 2011» [Электронный ресурс]. URL: аisori.meteo.ru/ClspR
12. Ребристая О.В. Флора полуострова Ямал. Современное состояние и история формирования. СПб.: Изд-во СПбГЭТУ «ЛЭТИ», 2013. 312 с.
13. Справочник по климату СССР. Л.: Гидрометиздат, вып.17, ч.I, II, 1965-1966, 276 с.
14. Телятников М.Ю.Особенности распределения тундровой растительности сибирского сектора Арктики. Диссертация на соискание ученой степени доктора биологических наук. Новосибирск, 2005. 320 с.
15. Хромов С.П., Мамонтова Л.И. Метеорологический словарь. Л: Гидрометиздат, 1974, 268с.
References:
2. Budyko, M. I. Climate and life. L., Hydrometeoizdat. 1981. Four hundred
3. Gashev S. N. Mammals of the Tyumen region: guide-determinant. Tyumen: Publishing house of TSU, 2008. 333 р.
4. Gelashvili D. B., Iudin D. I., Rosenberg G. S., etc. Fundamentals of the multifractal analysis of the species structure of the community // Successes of modern biology. 2008, vol. 128, Р. 21-34
5. Ilyina I. S., Lapshina E. I., Lavrenko N. N. and others. Vegetation cover of the West Siberian plain. Novosibirsk: Science, 1985. 250 p.
6. Konovalov A. A., Roman L. T. Features of Foundation design in oilfield areas of Western Siberia. Leningrad: Stroyizdat. 1981. 168 р.
7. Konovalov A. A., Gashev S. N., Kazantseva M. N. Distribution and hierarchy of biotic taxa in the territory of the Tyumen region // agrarian Russia. 2013, 4. P. 48-57.
8. Konovalov A. A., Glazunov, V. A., Moskovchenko D. V. Tigiev A. A. Gashev S. N. On the climatic dependence of the biota on the North of the Tyumen region. Materials of the III all-Russian meeting "Man and the North". Tyumen: publishing house of the Institute of Northern, 2015. P. 333-338.
9. Konovalov A. A., Ivanov S. N. Climate, phytoproduction and palinspastic: communication, distribution and methods of paleoreconstructions (on example of Western Siberia. Novosibirsk: GEO, 2007. 130 р.
10. Konovalov A. A. Climatic dependence of biota (on the example of Western Siberia). GOB, vol. 79, No. 1, 2018.Р. 55-63.
11. Scientific and applied reference book “Climate of Russia, 2011” [Electronic resource]. Mode of access: аisori.meteo.ru/ClspR
12. Rebristay O. V. flora of the Yamal Peninsula. Current state and history of formation. SPb.: Publishing house Etu "LETI", 2013. 312 р.
13. Guide to the climate of the USSR. Leningrad: Gidrometizdat, vol.17, CH. I, II, 1965-1966, 276 p.
14. Telaytnikov M/U. Features of distribution of tundra vegetation of the Siberian sector of the Arctic. Thesis for the degree of doctor of biological Sciences. Novosibirsk, 2005. 320 p .
15. Khromov S. P., Mamontova L. I. Meteorological dictionary. L: Hydromethysdate, 1974, 268 р.